На примере обыкновенной слепушонки Южного Урала установлено компенсаторное возрастание разнообразия в обедненных по составу популяциях. Показано, что в мономорфных популяциях разнообразие формы нижней челюсти выше, чем в полиморфных, где разделение кормодобывающих функций у зверьков разных морф приводит к общему снижению морфоразнообразия
Изучена морфологическая изменчивость формы нижней челюсти у моно- и полиморфных по окраске меха популяций обыкновенной слепушонки (Ellobius talpinus Pall., 1770) на Южном Урале. Для анализа использованы четыре географически удаленные популяции. Северная кунашакская (п. Кунашак, Челябинской обл.) и самая южная наурзумская (Наурзумский заповедник, Северный Казахстан) мономорфны по окраске, причем северные зверьки представлены исключительно меланистами, а южные – животными бурой окраски. Баймакская (п. Баймак, Республика Башкортостан) и троицкая (г. Троицк, Челябинская обл.) проявили полиморфизм окраски шкурки в виде трех дискретных морф: черная, бурая и чепрачная (широкая черная полоса на спине и бурое брюхо). Анализ формы нижней челюсти проведен по 15 меткам методами геометрической морфометрии (рис. 1а). Для оценки внутрипопуляционного морфологического разнообразия использован анализ паттерна ближайших соседних точек в пределах полигона изменчивости (Hammer et al., 2009).
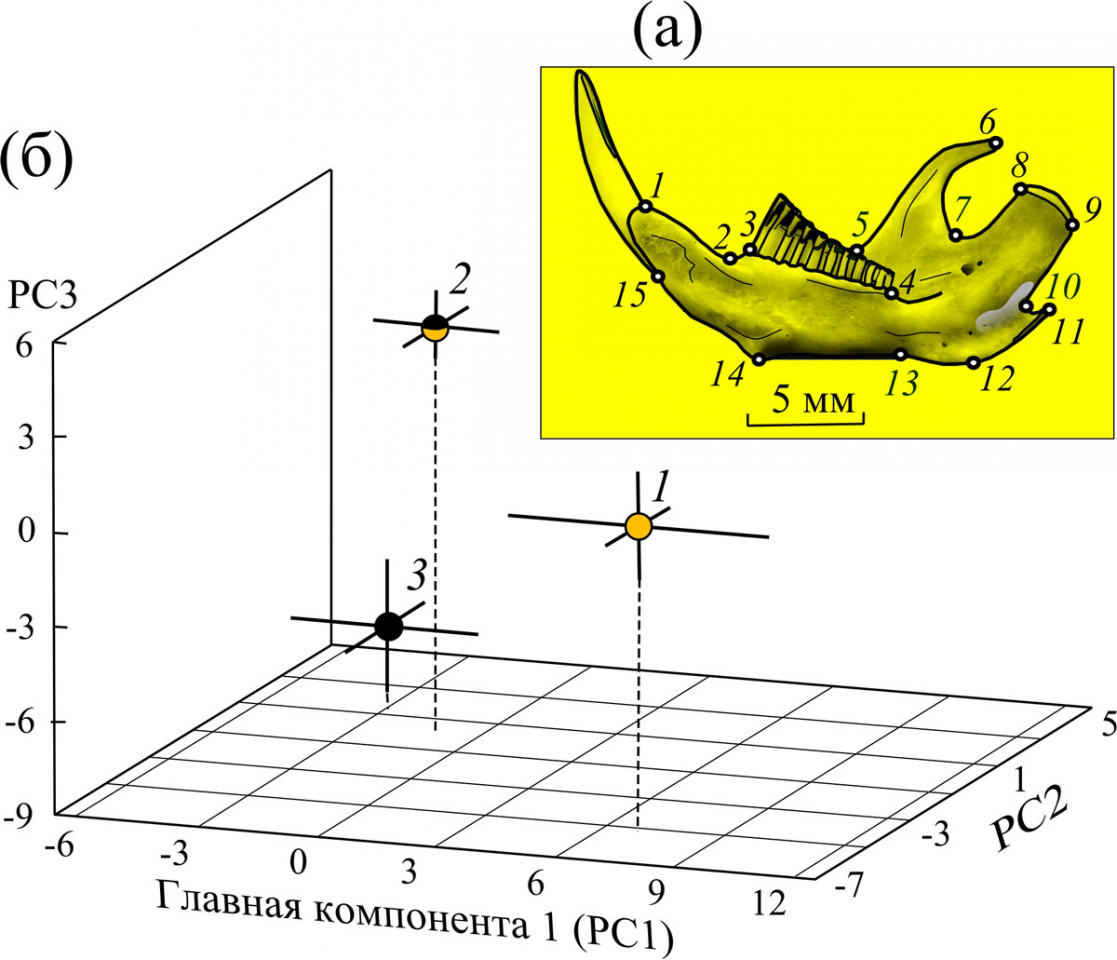
Рисунок 1 – Размещение (а) меток-ландмарок (1-15) на лингвальной стороне нижнечелюстной ветви обыкновенной слепушонки и (б) центроидов выборок трех морф по окраске шкурок (1 – бурые, 2 – чепрачные, 3 – черные) в общем морфопространстве вдоль первых трех главных компонент (PC1–PC3) формы нижней челюсти (с учетом стандартных ошибок – SE)
Установлено, что ординаты центроидов особей всех трех морф существенно разделены в морфопространстве первых трех главных компонент (рис. 1б). Отсутствие значимой ранговой корреляции Спирмена между размером центроида и первой главной компонентой (rsp= 0.08; p=0.444) исключает связь наблюдаемых различий формы с аллометрическими эффектами. Обнаруженная внутрипопуляционная дифференциация цветовых морф по форме нижней челюсти прямо указывает не только на их морфогенетические особенности, но и морфофункциональные различия, связанные с неодинаковой функцией мандибул зверьков разных морф при рытье и первичной обработке кормовых объектов. Результаты канонического анализа прокрустовых координат, характеризующих изменчивость формы нижней челюсти четырех сравниваемых выборок, представлены на рис. 2.
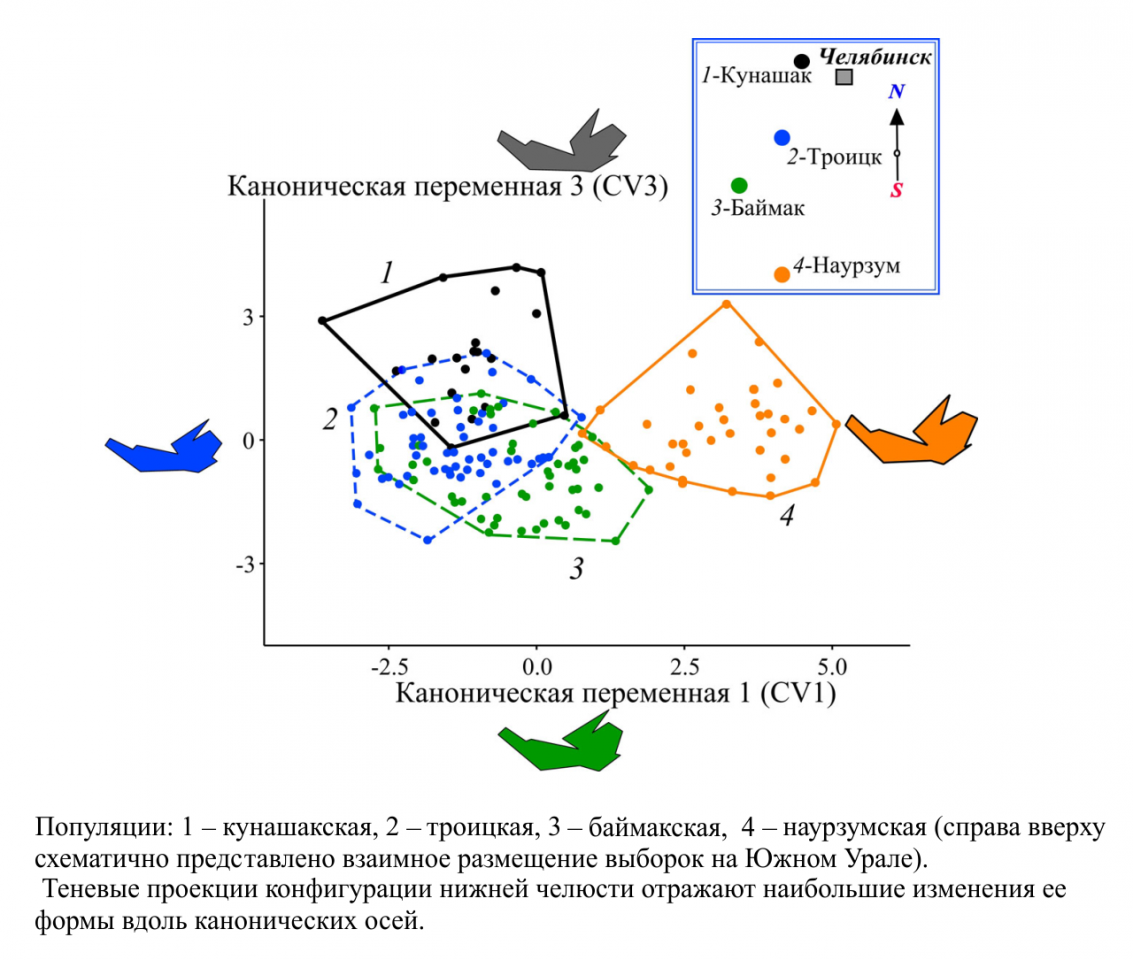
Рисунок 2 – Результаты канонического анализа Прокрустовых координат, характеризующих изменчивость формы нижней челюсти обыкновенной слепушонки
Межгрупповые различия вдоль всех трех канонических осей статистически значимы. Проявления межгрупповой изменчивости вдоль первой и третьей канонических переменных достоверно связаны с наличием в популяциях полиморфизма (r=-0.60; p<0.0001 и r=-0.48; p<0.0001 соответственно). Полигоны изменчивости обеих мономорфных популяций (наурзумской и кунашакской) разобщены и выборки существенно различаются по форме нижней челюсти. Полигоны полиморфных популяций (баймакской и троицкой) накладываются друг на друга в общем морфопространстве, но удалены от полигонов мономорфных группировок. Таким образом, форма нижней челюсти у мономорфных по окраске шкурки популяций (бурых и черных) различается, а у обеих полиморфных –сходна между собой. Типичная для мономорфных бурых зверьков конфигурация нижней челюсти отличается более массивным массетерным отделом и смещенным кверху сочленовным отростком, что указывает на ее высокий потенциал в осуществлении усилий, связанных с дроблением пищевых объектов (Anderson et al., 2014). Конфигурация мандибул мономорфных черных слепушонок имеет относительно суженное тело в области симфиза, значительное смещение венечного отростка вперед и вентральное отклонение сочленовного отростка, что свидетельствует о возможности приложения больших усилий в горизонтальном направлении при перетирании пищи. Полиморфные популяции имеют конфигурации, которые не позволяют потенциально достигать таких же усилий в вертикальном или горизонтальном направлениях, как у мономорфных. Сравнительная оценка внутрипопуляционного разнообразия формы нижней челюсти показала, что средние дистанции между ближайшими соседними ординатами (MNND) значимо выше у обеих мономорфных популяций по сравнению с полиморфными (рис. 3).
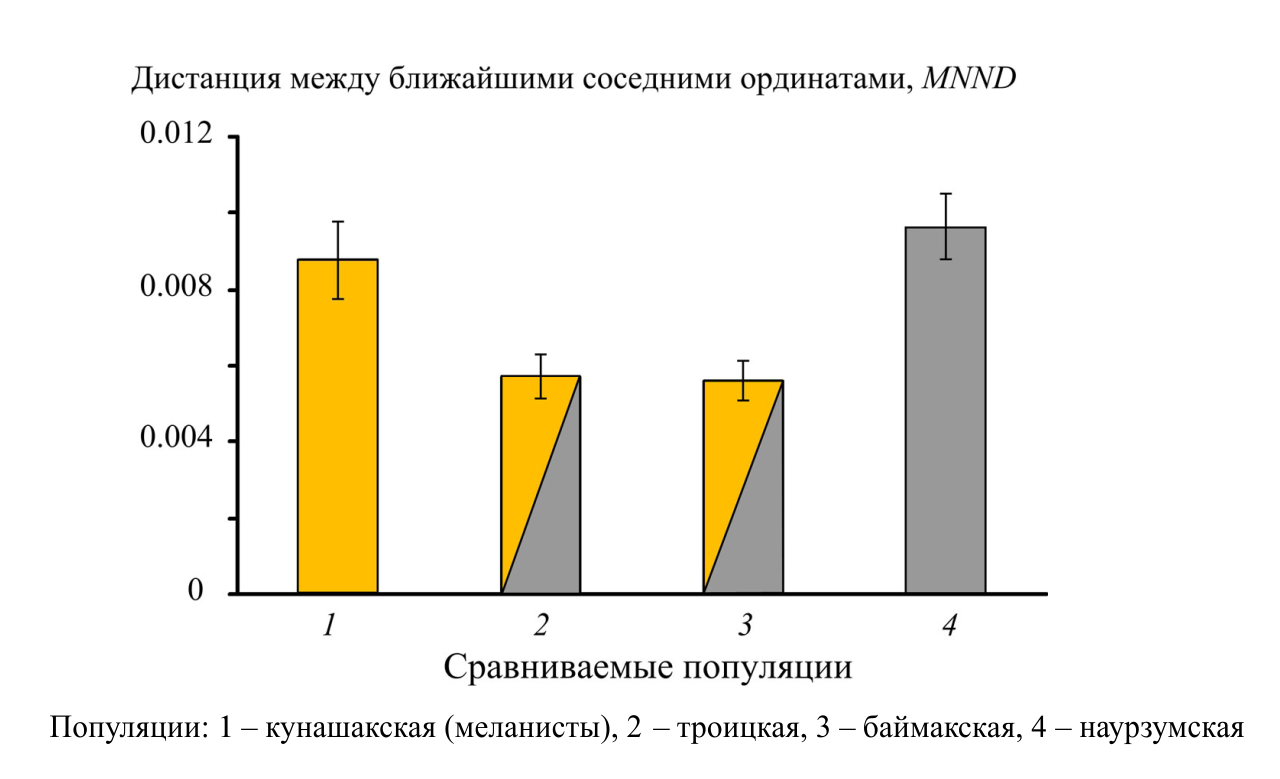
Рисунок 3 – Сравнение средних дистанций (MNND) с учетом стандартных ошибок (SE) между ближайшими соседними ординатами внутри выборочных полигонов изменчивости в мономорфных (1, 4) и полиморфных (2, 3) популяциях
Таким образом, в полиморфных популяциях в отличие от мономорфных наблюдается снижение морфоразнообразия нижней челюсти и формируется характерная конфигурация мандибулы, при которой относительно снижен потенциал усилий, доступных при дроблении и перетирании растительных объектов питания. Различия в форме нижней челюсти зверьков разных окрасочных морф отчетливо проявляются как при сравнении разных популяций, так и при сравнении зверьков разных морф внутри одной полиморфной популяции. Большее разнообразие формы мандибул в мономорфных популяциях по сравнению с полиморфными хорошо согласуется с принципом компенсации Ю.И. Чернова, что позволяет расширить область его применения на внутрипопуляционный уровень сравнений.