 На Общем собрании УрО РАН 4 декабря 2020 г. состоялось торжественное вручение медалей и дипломов Уральского отделения РАН. Медаль имени академика С.С. Шварца присуждена доктору биологических наук, заведующему лабораторией молекулярной экологии растений ИЭРиЖ Владимиру Леонидовичу Семерикову за цикл научных работ «Филогеография и эволюционная история деревьев бореальных и умеренных лесов».
На Общем собрании УрО РАН 4 декабря 2020 г. состоялось торжественное вручение медалей и дипломов Уральского отделения РАН. Медаль имени академика С.С. Шварца присуждена доктору биологических наук, заведующему лабораторией молекулярной экологии растений ИЭРиЖ Владимиру Леонидовичу Семерикову за цикл научных работ «Филогеография и эволюционная история деревьев бореальных и умеренных лесов».
Коллектив Института поздравляет Владимира Леонидовича с получением такой престижной и значимой награды. Публикуем интервью, в котором Владимир Леонидович рассказывает об истоках своей научной деятельности, о закономерностях расселения хвойных и широколиственных деревьев в Евразии и изучении их с помощью молекулярно-генетических методов.

Расскажите, почему вы пошли в науку? Как вы оказались в Институте? Ваш отец работал много лет в Институте, но далеко не всегда дети продолжают дело своих родителей, создавая научную династию.
Мой папа – Леонид Филатович Семериков – занимался наукой, все его друзья были из науки, разговоры дома были все время про науку. Поэтому, естественно, и я интересовался наукой всегда. В Институт экологии я попал сразу после окончания лесотехнического института.
У вас была интересная исследовательская жизнь, о каком периоде своей жизни вы вспоминаете чаще и скучаете – о жизни за рубежом или о работе на севере?
Пожалуй, и работа за рубежом была очень интересным и насыщенным в научном смысле периодом. Но, конечно, работа на севере, и, главное, походы на исследовательском судне «Флора» – это самая яркая часть моей жизни!
Чем именно вы там занимались?
В основном, конечно, занимался всем тем, что делает матрос на корабле: управлял судном, ставил паруса, прокладывал курс. Наблюдал за морем! Мы периодически останавливались для проведения исследований. В основном проводили сборы материалов по разным видам растений: по лиственнице, по кедру, по видам мятлика.
Походы на «Флоре», пожалуй, и были ключевыми моментами в моей научной карьере. Мы проводили сбор материала по лиственнице сибирской на северном пределе ареала. Ведь тогда была заложена глубокая идея – исследовать, каким образом вид способен колонизировать такие огромные пространства, причем с очень большим диапазоном условий. Лиственница растет на границе сухих степей на юге Башкирии и в Монголии, и в то же время обитает на Ямале, на Гыданском полуострове, на Таймыре. Такие широчайшие диапазоны способности к адаптации у таежных лесообразующих видов привлекают внимание людей, которые занимаются эволюцией, лесоведением и лесоводством, у лесников в особенности. Но чтобы это понять, надо вначале исследовать популяционную структуру вида. Мы исследовали популяционную структуру с помощью аллозимных методов. Тогда выяснились интересные вещи, связанные с географической структурой изменчивости.
Оказалось, что лиственница на севере состоит из двух отличных генетических групп – восточных популяций, которые обитают на Гыданском и Тазовском полуостровах, и группировки, которая обитает вдоль Урала и юго-западной части Ямала, в частности, на Хадыте. Это результат простого расселения. Первая группа расселялась из Сибири, а вторая – уральская. Они между собой смешиваются, но зона перехода довольно узкая, особенно на Ямале. На Хадыте в основном уральская популяция, а на Яда-Яходы-Яхе – чисто сибирская. Южнее, на основной территории Западной Сибири, эта полоса гораздо шире. Она начинается от Оби и вплоть до поселка Пангоды сохраняются гены и признаки обоих подвидов сибирской лиственницы. Причина этого лежит на поверхности, но чтобы это понять, нужно было начать исследовать генетику этой лиственницы. Имеется два источника распространения: горы Южной Сибири и Южный и Средний Урал. Собственно с этого и началась генетическая часть моей работы.

Парусно-моторная яхта "Флора" на р. Обь, 1991 г.
А чем вы занимались до работ на севере?
До этого я занимался биологической продуктивностью леса. Моим руководителем тогда был Смолоногов Евгений Павлович. Он был сотрудником Института, известным ученым, человеком широких интересов, которые были связаны в основном с экологией леса, с динамикой роста, взаимоотношениями пород и т.д. Он не касался генетики, но для меня работа с ним была большим опытом.
А потом возник интерес к генетике? Как это было?
Где-то в конце 80-х годов я заинтересовался генетикой, потом учился в аспирантуре в Москве. Там у меня была другая тема, связанная с сосной обыкновенной. Моим руководителем и другом моего отца – Львом Анатольевичем Животовским – была поставлена задача: можно ли с помощью имеющихся молекулярно-генетических методов (а тогда был только аллозимный анализ) определить, отличаются ли генетически популяции сосны обыкновенной, которые растут в хороших условиях и которые растут рядом на болоте. Болота есть везде на ареале сосны, и деревья на них всегда отличаются, имеют характерный вид. Собственно, с помощью аллозимного анализа я пытался понять, насколько генетически отличны плакорные и болотные формы сосны. Хотя и наивно было надеяться, что мы что-то сумеем определить. Геном у сосны обыкновенной огромный, а аллозимы – это всего несколько доступных для анализа локусов. Было выяснено, что аллозимы не показывают различий между суходольной и болотной соснами. Но Станислав Николаевич Санников и Ирина Владимировна Петрова показали, что эти формы различаются и имеют наследуемые адаптивные признаки. Не совсем понятно, генетические ли это признаки. Ведь любой организм имеет, кроме генетики, и сферу эпигенетики, которая определяет приобретенные признаки. Часть из них тоже наследуется, переходит из поколения в поколение. Хотя эти изменения не затрагивают собственно геном и нуклеотидные последовательности.


Владимир Леонидович Семериков и Вадим Борисович Федоров в лаборатории Университета города Уппсала, 1999 г.
Медаль Шварца вы получили за многолетнюю работу по изучению филогеографии и эволюционной истории деревьев бореальных и умеренных лесов. Расскажите подробнее о своих исследованиях. Чем вы сейчас занимаетесь?
А сейчас я чем только не занимаюсь! В основном, конечно, мы работаем с филогеографией разных видов растений – хвойных, лиственных. Опять мы вернулись к дубу, к тому, с чем мой папа был связан, но мы продолжили на другом уровне. Этим в основном занимается Светлана Александровна Семерикова.
Расскажите, пожалуйста, что происходит с дубом?
Это новый виток нашей работы: изучаем популяционную структуру дуба, в том числе на Северном Кавказе и в Европейской части России. В 90-х годах были предприняты исследования популяционной структуры широколиственных европейских видов. По маркерам хлоропластной ДНК было установлено, что наибольшая изменчивость европейских видов – дуба черешчатого, скального, пушистого, липы, бука, ясеня и других широколиственных находится на юге Западной Европы, в районе побережья Средиземного моря. И чем дальше на север, тем генетическая изменчивость все меньше. При этом гаплотипы, которые есть в районе Средиземного моря, встречаются и на севере, но часть из них теряются по пути. И по распространению отдельных гаплотипов можно проследить пути их расселения. Если смотреть на их географическое распространение, можно увидеть полосы или шлейфы, направленные с юго-запада на северо-восток. Это указывает на то, что виды распространялись из отдельных регионов Средиземноморья – Испании, Италии, Балкан – на северо-восток по территории Западной Европы. Был сделан вывод, что широколиственные виды, такие как дуб, распространялись совсем недавно, после последнего ледникового максимума, во время которого дуб сохранялся только в очень теплых средиземноморских убежищах. Такая парадигма была распространена на весь ареал дуба, липы и других видов широколиственных деревьев.
Однако, на восточном крае ареала этих видов ситуация совершенно не похожа на то, что было выявлено в Западной Европе. Это также подтверждается исследованиями морфологических признаков того же дуба. По хлоропластным маркерам в Западной Европе и у нас (в России) оказалось не похоже, что дуб пришел с запада в голоцене. Вероятно, он здесь сохранялся в последний ледниковый максимум, и, возможно, даже пережил здесь несколько ледниковых фаз. Расселялся из каких-то местных источников с Южного Урала или из Среднего Поволжья. Мы обнаружили, что на Русской равнине имеется резкая граница по линии Смоленск–Псков–Санкт-Петербург. Все, что западнее этой линии, имеет высокую изменчивость, много гаплотипов у дуба и у липы, а восточнее (на линии Москвы точно уже) остается всего два гаплотипа у дуба черешчатого и два у липы. Причем этих гаплотипов нет западнее (в Белоруссии, Западной Украине, Ленинградской, Псковской областях и т.д.), там другие гаплотипы. Таким образом, они здесь давно существуют, когда-то, может миллион лет назад, в плейстоцене, они сюда попали из Европы или с Кавказа и здесь осели и не исчезали насовсем никогда. Ареалы дуба и липы сжимались каждый раз до маленьких популяций, потом снова расширялись.
У липы история даже еще интереснее. Липа сибирская в Кемеровской области и Алтайском крае заметно отличается от нашей – европейской мелколистной липы Tilia cordata. Исследовали это, в том числе, и англичане (недавно была работа), и по ядерным маркерам оказалось, что липа сибирская Tilia sibirica образует совсем другую группировку, отдельную ветвь. Также был оценен возраст. С Tilia cordata они разошлись где-то в среднем плейстоцене. Но эти исследователи упустили, что липа сибирская есть не только в Кемеровской области, еще существует два маленьких локалитета в Красноярском крае. И красноярские коллеги с нашей подачи их проанализировали и обнаружили, что там вообще другая липа. Она не похожа на кемеровскую популяцию, но отличается и от европейской липы. Отличия хоть и небольшие, но явные. Причем по хлоропластной ДНК кемеровская сибирская липа имеет гаплотип, который распространен как в Сибири, так и в Восточной Европе, вплоть до Восточной Белоруссии. Этот гаплотип очень широко распространен и у нас на Урале, в Поволжье, в Подмосковье, на юг она ограничена районом южных гор – Кузнецкого Алатау и Алтая. В общем, сложная была история взаимоотношений липы сибирской и европейской.
Видимо липа сибирская, к которой относится кемеровская популяция, была распространена очень широко, и ее гаплотип закрепился на Русской равнине. А потом, когда настал голоцен и стало тепло, более теплолюбивая европейская липа вытеснила сибирскую, но не физически, а в процессе интрогрессивной гибридизации, путем опыления. В новых условиях она более сильная, жизнеспособная, производит больше пыльцы и семян. В итоге европейская липа заместила сибирскую до самого Иртыша, но хлоропласты остались прежними – от сибирской. Хлоропласты передаются по материнской линии, они не могут передаваться с пыльцой. А в Красноярском крае, вдоль Енисея, вероятно, вообще третий вид. Он когда-то расселился с запада из Европы и ограничился этим районом. Интересно, что этот красноярский гаплотип встречен еще только в Ленинградской области. То есть в пространстве между Ленинградской областью и Красноярском после расселения он был полностью вытеснен другими гаплотипами.
Сбор материала, 2020 г.
Ваша публикация об истории расселения пихты в Евразии попала в доклад о важнейших достижениях российских ученых в 2018 году. Расскажите, пожалуйста, об этом более популярно.
Да, конечно! Мы занимаемся филогенией пихт, исследуем, откуда произошли современные виды пихт, какие виды от каких, когда возникло их разнообразие. Для этого используем молекулярные маркеры, хлоропластные, митохондриальные, ядерные маркеры ДНК и палеонтологические данные. Мы секвенировали на нашем секвенаторе в большом объеме ядерные фрагменты ДНК пихт. В результате исследования этой филогении оказалось, что современные виды пихт диверсифицировались, то есть первично распространились, весьма недавно – менее 20 млн. лет назад. Хотя остатки пихт встречаются гораздо раньше – 40–45 млн. лет назад, а пыльца пихт широко отмечается и в мелу. Древние виды пихт вымерли, а современные возникли снова от какой-то одной уцелевшей формы. И наиболее дивергентные по ядерным и хлоропластным маркерам формы пихт, то есть виды, которые сильно отличаются друг от друга и всех остальных, находятся в Америке. То есть изначальная диверсификация современных пихт началась в Америке, а потом уже через какое-то время произошло их расселение в Северную Евразию. Причем этих вселений в Евразию было несколько, и каждое последующее как бы стирало предыдущее разнообразие, замещало предыдущие формы.
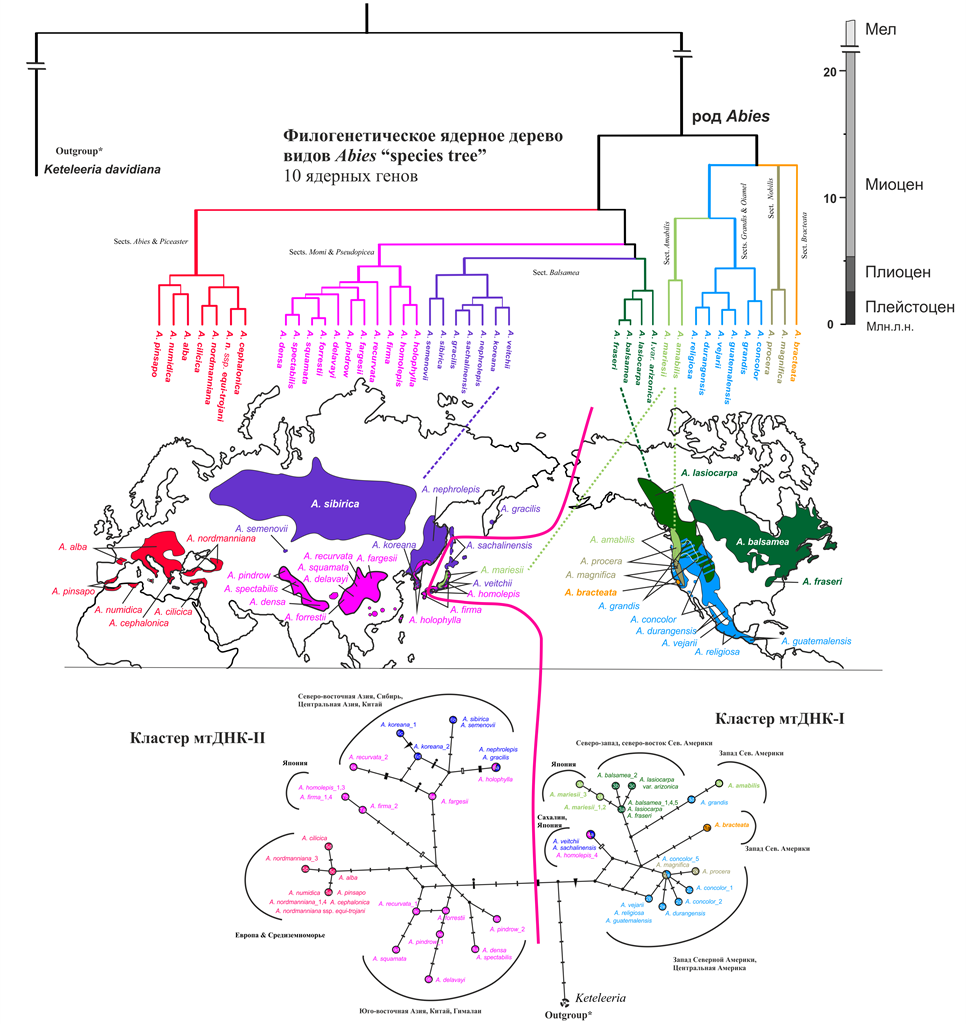
Филогения пихт по ядерной и митохондриальной ДНК, иллюстрация из статьи в журнале Molecular Phylogenetics and Evolution. 2018
За счет чего это могло произойти? Какие механизмы заложены в процессе расселения одних видов и вытеснения других?
Вероятно, каждые новые эмигранты из Америки были более адаптивны к новым условиям. Эволюции в Америке шла более эффективно, чем в Евразии. В Америке были довольно большие территории с холодными условиями, пригодные для обитания и эволюции бореальных видов, а в Евразии длительное время были вообще тропики вплоть до самого севера, и после тоже довольно длительное время были достаточно мягкие условия, то есть мест для эволюции таких хвойных было меньше. Виды приходили в Евразию и оставались без особых изменений, а в Америке они быстро совершенствовались. Возможно, это еще было связано с тем, что каждый вид в ходе своей длительной эволюции обрастает множеством паразитов и болезней, к которым он приспосабливается. Например, имеются два родственных вида, которые отделены друг от друга, у каждого свои болезни, и при встрече они обмениваются. И часто этот обмен выглядит так, что один из видов исчезает, не выдержав чужих патогенов. Также ведь и инвазивные виды проникают, причем они оказываются часто невосприимчивыми к местным патогенам и инфекциям, но сами что-то приносят, что уничтожает местные виды. Например, у нас в Сибири уже много столетий происходит вытеснение сибирской лиственницы даурской. Мы видим, что раньше она встречалась шире, ее генетические следы проявляются гораздо восточнее. Например, в устье Хатанги в лиственничных редколесьях встречаются признаки как сибирской, так и даурской лиственницы. Там они даже морфологически сходны, но по ядерным маркерам они в основном даурские. При этом даурская лиственница идет на запад вплоть до Норильска, почти до Енисея распространена. Когда-то сибирская лиственница доходила до устья Лены, а сейчас даурская ее вытеснила. И на юге происходит примерно то же самое. Такие же процессы происходили и раньше, когда одни виды вытесняли другие...
Очень интересно! А что в итоге произошло с пихтами?
Да, вернемся к пихтам. По-видимому, было несколько волн миграций. От самой-самой ранней волны миграции из Северной Америки остались средиземноморские виды. Эти виды пихт прошли через всю Евразию и осели вокруг Средиземного моря. Они сохранились там, потому что оказались слишком далеко от входа, от Берингова пролива. Они оказались там изолированными. Потом была вторая волна, которая принесла южно-азиатские виды, ее следы сохранились в Гималаях, на юге Китая, и некоторые виды в Японии несут черты этих ранних миграций. Потом была следующая волна, она принесла бореальные виды пихт, такие как наша сибирская и дальневосточные виды – белокорая, сахалинская и камчатская. Последние две ближе всего к американским видам. У них митохондриальная ДНК сохранилась от американских видов, а хлоропластная ДНК уже азиатская, ядерная – наполовину американская. И самый американский вид на территории Северной Евразии – это пихта Мариса Abies mariesi, это уже японский вид. В Японии 5 видов пихт, и один из них (пихта Мариса) практически один в один совпадает с одним из американских видов – пихтой миловидной Abies amabils, даже морфологически похож. Хотя они живут на противоположных сторонах Тихого океана, но генетически очень близки. Значит A. amablis совсем недавно прошла вдоль тихоокеанской дуги с востока на запад, перешла Берингов пролив, зашла в Японию и осела тут. Вот такие интересные вещи.
Хотя и там остались нерешенные задачи и проблемы. И нам понадобилось исследовать филогению митохондриальной ДНК пихт, хоть и исследовали ее ранее и там очень низкая изменчивость. Митохондриальная ДНК у растений очень медленно эволюционирует, имеет низкую скорость мутирования, и поэтому информативной изменчивости, которая может быть использована в филогенетических построениях, там мало. Чтобы что-то уловить, необходимо секвенировать очень большие объемы. К сожалению, наши методы, которые используются чаще всего, – секвенирование по Сэнгеру с помощью капиллярного электрофореза – не дают нужного разрешения. И сейчас у нас проект, который идет второй год, включает изучение филогении митохондриальной ДНК пихт на расширенном материале. Мы использовали для этого секвенирование следующего поколения с помощью прибора Illumina. Этот метод используется сейчас для секвенирования целых геномов, изучения метагеномики, целых сообществ грибов, бактерий. Хоть это и очень мощный метод, но целиком геном хвойного растения эта машина секвенировать не может. У хвойных просто гигантский геном. Если у человека геном, например, 3 гигабайта, то у пихты – около 20 гигабайт. И поэтому секвенирование полного генома хвойных очень большая и трудная задача. Сейчас сделано всего несколько экземпляров. Но для исследования изменчивости нужно же не один геном отдельного дерева, необходимо секвенировать большую выборку. Метод полного секвенирования для изучения изменчивости не подходит, поэтому надо брать только митохондриальные ДНК. Мы используем такой подход, называется гибридный захват. С помощью ПЦР создаем зонды митохондриальной ДНК какого-то хвойного дерева, например лиственницы сибирской. Зонды в данном случае это участки митохондриальной ДНК. Их помещаем в раствор в пробирку, туда же ДНК конкретной особи, которую хотим исследовать. Нагреваем и выдерживаем 2-3 суток при пограничной температуре (68° С), при которой происходит реассоциация ДНК, то есть разделение на двуцепочечную. При этом гомологичные участки начинают склеиваться, и зонды садятся на ДНК нужного участка. Зонды подготовлены специальным образом – к ним добавлена молекула биотина, это позволяет извлекать их с помощью магнитных частиц, которые покрыты стрептавидином (биотин и стрептавидин при встрече образует ковалентную связь – это свойство широко используется в молекулярке). Потом все помещаем в магнит, магнитные частицы прилипают к магнитным стенкам, все остальное удаляем, промываем, нагреваем, смываем нужные участки ДНК и секвенируем уже их с помощью Иллюмины. В итоге мы получили очень большие данные, которые надо обрабатывать. Я, к сожалению, не владею биоинформатическими методами. У нас есть коллаборатор – Путинцева Юлия из Красноярска. Она имеет опыт, занималась секвенированием генома лиственницы. И сейчас она занимается обработкой наших данных, которые вышли очень объемными. Были проблемы даже со скачиванием файлов. Но работа идет на полных парах.
Над какими еще проектами вы сейчас работаете?
Еще у нас идет работа по исследованию генетической и эпигенетической составляющей адаптации хвойных растений к климату. Я в этой работе хоть и руководитель, но сильно влиять на ее ход не могу, потому что основные исполнители находятся в Красноярске, в геномном центре. Лиственница сибирская продолжает привлекать исследователей. Хотим выявить генетическую, и параллельно, эпигенетическую основу адаптации от юга до севера у лиственницы сибирской и лиственницы Сукачева. Лиственница Сукачева это западный подвид сибирской. Это как бы два параллельных вида, не связанных между собой, и если у них будет общий тренд адаптации с юга на север, то это и есть то, что мы ищем. Мы до сих пор не знаем, что лежит в основе этой адаптации. Возможно, у неё генетическая природа.
В процессе расселения с юга на север отбираются некие аллели разных генов, их комбинации приводят к тому, что вид существует в разных условиях. В условиях севера – короткого вегетационного сезона, но при этом длинного дня – вид способен вовремя прекращать рост, закладывать почки и, наоборот, выходить в вегетацию. И в то же время вид адаптирован к югу, даже к нашей широте. Если мы возьмем лиственницу с севера и посадим у нас, то она прорастет, какое-то время будет расти, а потом окажется «в раздумьях», что ей делать? Для нее день уже короткий. На севере она растет при почти круглосуточном освещении, а у нас уже 5-6 часов темноты даже в конце июня. Тогда она сразу прекратит свой рост и впадет в покой. То есть это дерево у нас не сможет расти в принципе. А если мы, наоборот, семена отсюда посадим, например, в Лабытнангах, то дерево будет усиленно расти до августа, когда все деревья уже начинают желтеть. Они уже два месяца как прекратили рост, а наша – растет. И с наступлением зимы и мороза эта лиственница просто замерзнет и погибнет. Таким образом, местные генотипы адаптированы к своему местообитанию, но все-таки непонятно, генотип это или эпигенотип.
Возможно, причина и не в том, что в генах есть какой-то нуклеотидный полиморфизм, а в эпигенетической адаптации. Эпигенетика в целом играет очень важную роль в развитии организма. Из хвои никогда не получится корень, и, наоборот, из корня – хвоя. Закладываются определенные настройки в клетках, которые направляют развитие и у растений и у животных. Один из путей – это метилирование цитозина в каких-то участках ДНК. Если это происходит в промоторе гена, который кодирует какой-то белок начала транскрипции, то подавляется синтез матричной РНК при транскрипции. Так происходит настройка этого гена, чтобы его не было слишком много, то есть организм сам подкручивает настройки. А одна из гипотез, что некоторые эпигенетические настройки наследуются. И, возможно, какие-то из этих изменений позволяют следующему поколению уже знать, когда будет весна или осень, когда вступать в вегетацию, когда время начинать цветение и заканчивать его. И мы сейчас это исследуем, хоть и возникают методические трудности в лабораторном плане. Но надеемся, что все получится!
Какие публикации вы готовите в настоящее время?
Мы сейчас продолжаем цикл публикаций, и про пихты в том числе. Пока небольшие, но для больших статей еще не пришло время. Все впереди!
И последний вопрос. Чего бы вам хотелось пожелать коллегам?
Коллегам можно пожелать сохранения интереса к науке! Когда у человека есть интерес к науке, он работает. Еще, естественно, грантов, проектов, удачи в публикациях. Чтобы рецензенты были не столь строги. И здоровья, конечно!

Решение о присуждении медалей и дипломов Уральского отделения РАН с премиями для сотрудников организаций академической, вузовской и отраслевой науки, находящихся под научно-методическим руководством УрО РАН опубликовано в постановлении президиума УрО РАН от 17.09.2020. Медаль имени С.С. Шварца вручается за научные труды, научные открытия и изобретения, имеющие большое значение для науки и практики в области биологических наук.
Беседовала Ю.В. Городилова
Фотографии и иллюстрации предоставлены В.Л. Семериковым