Обобщены многолетние данные по экологии личинок пеляди в нерестовых притоках нижней Оби; показана межгодовая вариабельность питания и условий среды во время их перехода на экзогенную пищу. К ключевым условиям, которые решают судьбу поколения, относятся как биотические (концентрация кормовых организмов), так и абиотические (ветровой режим) факторы. Оптимум трофического фактора во время перехода личинок пеляди на потребление пищи извне, обеспечивающий достаточную для выживания и роста накормленность в условиях пойменного водоема, определяется комплексом природных событий
С целью определения оптимума трофического фактора во время перехода личинок обской речной пеляди на активное питание проанализированы данные, собранные в пойменном водоеме нерестового притока нижней Оби (сор Польхостур) в 1985–1999 гг. Установлено, что условия, при которых проходит миграция личинок пеляди по реке, складываются разнообразно как в разные годы, так и в течение одного сезона, что, наряду с разнокачественностью во время вылупления, определяет неоднородность по величине энергетических запасов. Покатные личинки пеляди подходят к сору Польхостур с остаточным запасом желтка или без него, но у всех особей сохраняется жировая капля. Большинство генераций пеляди переходят на экзогенное питание с небольшими запасами эндогенной пищи, а в 36.4% эти запасы отсутствовали. В некоторой степени этому способствовало сравнительно высокое (выше 10°С) прогревание воды в соре (r=–0.552).
Доля личинок пеляди в соре, растративших запасы энергетических ресурсов, но не перешедших на потребление корма извне, сильно варьировала в разные годы: в большинстве лет (66.7%) она составляла от 10.0% до 25.0%, в остальные годы – от 37.5% до 72.0%. В пойменном водоеме в большинстве лет переход на экзогенное питание личинок пеляди проходил при низкой концентрации кормовых организмов (рис. 1). В годы высокой смертности личинок (выше 95.0%) концентрация их стартового корма на мелководьях сора не превышала 8 тыс. экз./м3, поэтому эту величину мы принимаем как пороговую.
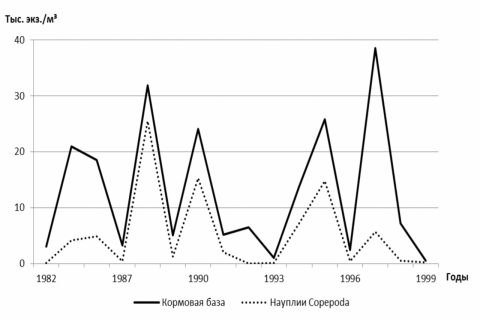
Рисунок 1 – Численность кормовой базы и излюбленного корма личинок пеляди во время перехода на экзогенное питание на мелководьях сора Польхостур
Накормленность генераций пеляди в соре находится в прямой зависимости от уровня развития кормовой базы: от совокупности кормовых организмов (r = 0.770); от науплиусов веслоногих рачков (r = 0.653); от меропланктона (r = 0.576); от типичных зоопланктеров (r = 0.761). Кроме концентрации стартового корма, накормленность ранних личинок пеляди в соре Польхостур определяют и абиотические факторы, влияние которых может быть как непосредственное, так и косвенное. Так, накормленность ранних личинок пеляди в соре была выше при западном, северо-западном и северном ветрах (r = 0.860) и ниже – при южном, юго-восточном и восточном (r = –0.835). При западном, северо-западном и северном ветрах на подветренных биотопах с благоприятными концентрациями кормовых организмов личинки имеют возможность спокойно потреблять пищу.
Температура воды в соре Польхостур во время подхода ранних личинок пеляди к мелководьям во все годы исследований превосходила низкую и не превышала верхнюю пороговые величины, при которых ранние личинки пеляди могут питаться, и в 60% лет ее значения лежали в интервале 8–11оС (рис. 2). Зависимости накормленности и смертности личинок пеляди от температуры воды во время перехода на активное питание не выявлено. Косвенное влияние температурных условий водоема на выживаемость личинок сиговых (например, через развитие кормовой базы) достаточно велико, но маловероятно, что в природных условиях температура воды может быть непосредственной причиной гибели личинок сиговых рыб. Подтверждает это отрицательная корреляция накормленности генераций пеляди и темпа прогревания воды в соре (r = –0.752). Такая связь опосредованно отражает уровень развития кормовой базы в разные годы, т.к. при быстром прогревании воды в соре наблюдали высокий темп развития рачкового планктона.
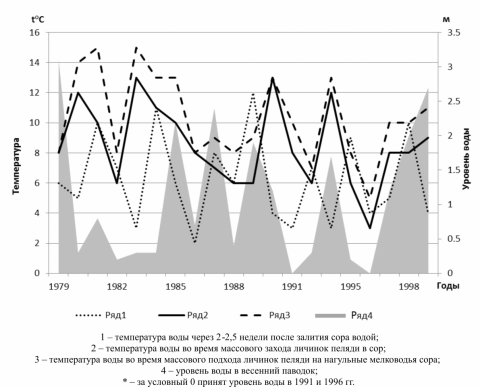
Рисунок 2 – Температура и уровень воды* в соре Полхостур в весеннее время
Таким образом, показано, что в соре Польхостур трофический фактор личинок пеляди во время перехода на экзогенное питание отличается значительной межгодовой вариабельностью и в отдельные годы определяет смертность генерации этого вида, которая изменяется с 65.1 до 98.2% (Богданов, Богданова, 2010). Непосредственно препятствовать переходу ранних личинок пеляди на активное питание могут концентрация кормовых организмов ниже 8 тыс. экз/м3 и неблагоприятный ветровой режим, способствующий образованию и накату волн на наиболее благоприятные нагульные биотопы, что, наряду с прямой гибелью личинок, снижает доступность кормовых организмов. Опосредованно, через развитие кормовой базы и увеличение волнения воды, такими факторами могут быть температурный и гидрологический (уровень весеннего паводка) режимы водоема. Наиболее сложная трофическая ситуация для ранних личинок пеляди складывается в те годы, когда совпадает позднее весеннее половодье и ранний скат личинок; когда при раннем залитии сора заход в него личинок проходит в поздние сроки, при этом усугубить ситуацию может высокий темп прогревания воды, который приведет к «перерастанию» кормовых организмов, прежде всего, ветвистоусых рачков.