С использованием микросателлитных маркеров ядерной ДНК в импактных ценопопуляциях горицвета (Lychnis flos-cuculi), произрастающих на территориях, сильно загрязненных промышленными выбросами, выявлен эффект основателя в отдаленном прошлом и повышение ауткроссинга в последних поколениях. Установлено, что самоопыление у этого вида не служит механизмом фиксации аллелей устойчивости к загрязнению и изоляции от неустойчивых ценопопуляций
Результаты анализа факторов, определяющих специфическую генетическую структуру ценопопуляций растений загрязненных местообитаний, противоречивы, что затрудняет их обобщение и требует дальнейшего накопления информации. Особый интерес представляют оценки вклада самоопыления в генетическую изменчивость у видов со смешанным типом опыления.
В трех зонах загрязнения выбросами Среднеуральского медеплавильного завода (в 30 км от завода на фоновой территории, в 4 км – на умеренно загрязненных участках и в 1.5 км – в сильно загрязненных местообитаниях) в девяти ценопопуляциях проведен сбор 1059 семян, а также образцов листьев и стеблей со 135 взрослых особей L. flos-cuculi – лугового многолетнего поликарпика со смешанным типом опыления – для последующего генотипирования с помощью шести микросателлитных маркеров ядерной ДНК.
Доли гетерозигот (Рис. 1А, Б) и аллельное разнообразие (Рис. 1В) в ценопопуляциях сильно загрязненной зоны ниже, чем на буферной и фоновой территориях, причем как в родительском поколении, так и в потомстве. Это свидетельствует об эффекте основателя в ходе заселения местообитаний вблизи завода или в результате элиминации значительной части ценопопуляций, возможно, из-за токсического стресса. По результатам анализа межпопуляционной генетической изменчивости, максимальные значения попарных FST (0.232) обнаружены между ценопопуляциями сильно загрязненной и остальных территорий, а также межу ценопопуляциями сильно загрязненной территории (0.040–0.096), тогда как Fst между ценопопуляциями фоновых и умеренно загрязненных участков не превышали 0.042. Такая значительная генетическая дифференциация ценопопуляций сильно загрязненной территории – следствие эффекта основателя.
Разница (ΔFIS) между FIS родительского поколения и потомства в большинстве ценопопуляций отрицательна (Рис. 1Г), что свидетельствует о пониженной жизнеспособности инбредных особей, а это согласуется с ранее полученными результатами экспериментальных исследований L. flos-cuculi. При этом ΔFIS в ценопопуляции из максимально загрязненного местообитания положительна, что указывает на повышенную частоту ауткроссинга в последнем поколении. Кроме того, по результатам анализа избытка гетерозигот, выполненного с помощью ПО Bottleneck, для ценопопуляции максимально загрязненного участка выявлены признаки популяционного взрыва в последних поколениях (при всех моделях мутирования генетических маркеров получены p≤0.03 в пользу гипотезы о дефиците гетерозигот). Полученные результаты могут быть хорошо обоснованы с точки зрения временной динамики условий среды в исследуемых местообитаниях: под влиянием загрязнения на участках вблизи источника выбросов происходит изреживание лесных массивов, что приводит к постепенному увеличению площади подходящих для L. flos-cuculi местообитаний и снижению барьеров для переноса пыльцы и семян между соседними ценопопуляциями.

Рисунок 1 – Наблюдаемая (А) и ожидаемая (Б) частоты гетерозигот; аллельное разнообразие (В) и коэффициент инбридинга (Г) в ценопопуляциях L. flos-cuculi
Расчет частоты самоопыления (S) в исследованных ценопопуляциях проведен на основе данных по генотипированию двух поколений вручную, а также с помощью четырех методов, реализованных в программах RMES, MLTR, COLONY2. В основе этих методов лежит предположение о том, что если в генотипе проростка обнаружены аллели, отличные от аллелей материнского растения, то они были принесены с пыльцой чужеродного отцовского растения, отсутствие отличий – обусловлено самоопылением. Различия между методами заключаются в подходе к оценкам вклада мутаций исследуемых маркеров и скрещивания близкородственных генотипов в S.
Оценки S, полученные c помощью разных методов, различаются. В частности, частоты самоопыления, рассчитанные в RMES, во всех ценопопуляциях выше по сравнению с результатами других методов, тогда как при использовании COLONY (метод FPLS), S завышены в ценопопуляциях с высоким генетическим разнообразием. Для ценопопуляций с большим количеством клонов (№2 и №3 в сильно загрязненной зоне) S, полученные разными методами, различаются кардинально (от 0–0.1, полученных в COLONY 2 до 0.5–0.8 – в остальных). Между тем, для ценопопуляций с высоким генетическим разнообразием (все ценопопуляции фоновой и умеренно загрязненной зон, а также ценопопуляции № 1 из сильно загрязненной зоны) S, полученные вручную, c помощью MLTR и COLONY (метод PLS) согласуются. Они составляют 0.07–0.23 и не различаются значимо между зонами загрязнения (рис. 2).
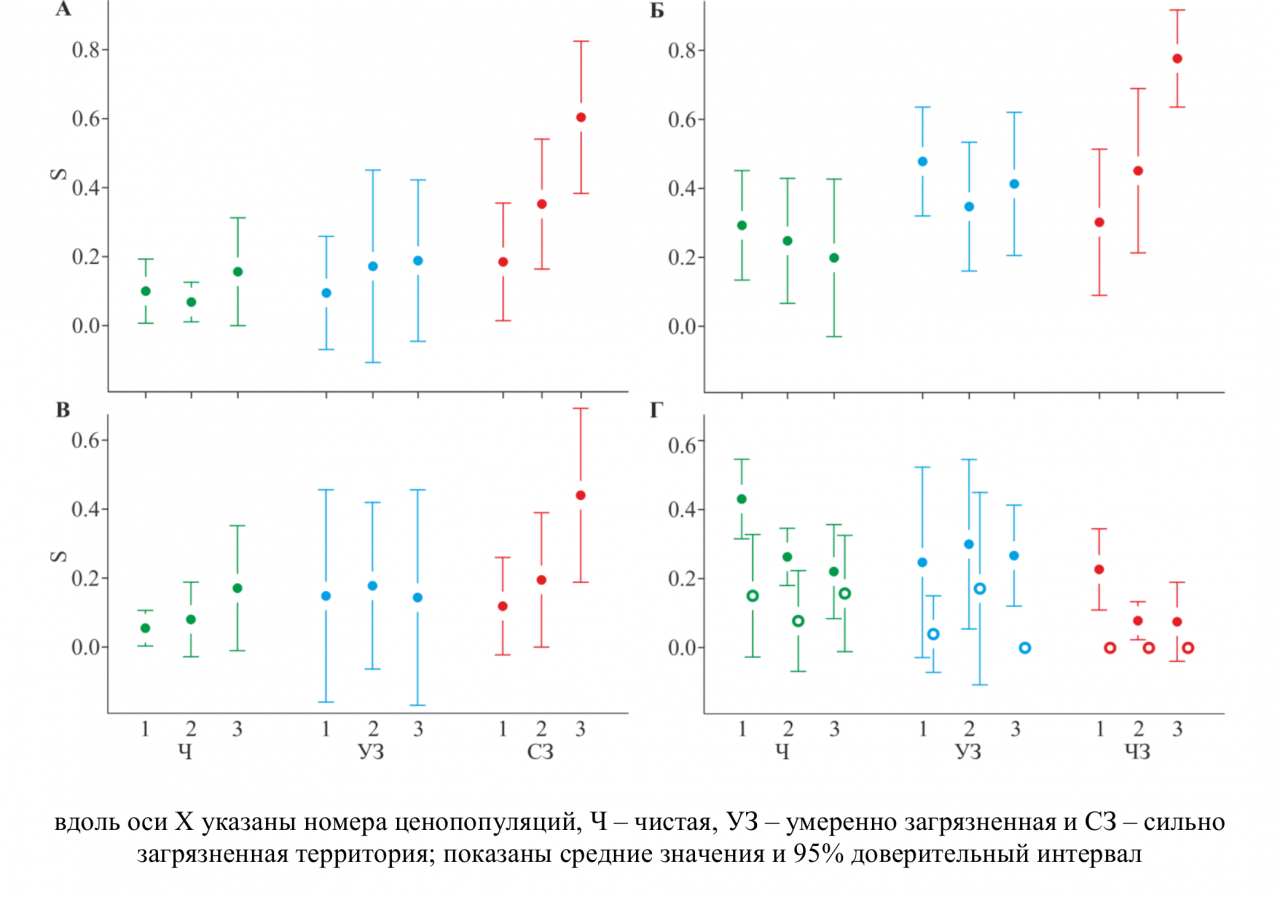
Рисунок 2 – Частота самоопыления (S) в ценопопуляциях L. flos-cuculi, рассчитанная вручную (А) и с помощью RMES (Б), MLTR (В), COLONY2 (Г) методом PLS (закрашенные круги) и FPLS (пустые круги)
Полученные результаты свидетельствуют о том, что вклад самоопыления в семенное размножение у L. flos-cuculi незначителен. При этом самоопыление не служит механизмом фиксации аллелей устойчивости к загрязнению или репродуктивным барьером для скрещивания с соседними неустойчивыми популяциями, вопреки результатам, полученным для ряда других видов травянистых растений из промышленно загрязненных зон. Важный методический вывод заключается в том, что при расчете частот самоопыления обязательно использование разных методов анализа во избежание ошибочных заключений, хотя это и не гарантирует получения однозначных выводов.