Анализ изменчивости фрагментов хлоропластной ДНК группы близкородственных рододендронов подсекции Rhodorastra со спорным таксономическим статусом (Rhododendron dauricum s.s., Rh. ledebourii, Rh. sichotense и Rh. mucronulatum; Ericaceae) выявил высокую генетическую дифференциацию в ее пределах, подтверждая таксономическую самостоятельность указанных видов. Получены оценки параметров внутрипопуляционной генетической изменчивости, выявлены особенности биогеографии данных видов
Изучен комплекс викарирующих близкородственных рододендронов со схожей морфологией и запутанной систематикой – рододендроны Ледебура (Rh. ledebourii Pojark.), даурский (Rh. dauricum L.), сихотинский (Rh. sichotense Pojark.) и остроконечный (Rh. mucronulatum Turcz.). Ареалы данных видов занимают территорию от Алтая до Приморья. В работе проведено сравнение популяционной структуры, особенностей генетической изменчивости, путей эволюционных преобразований и биогеографии этих видов на основе данных изменчивости хлоропалстной ДНК.
Анализировали 4 полиморфных фрагмента: TabCD, TabAB, участок matK и CS (названия фрагментов приводятся в соответствии с Rendell, Ennos, 2002). По каждому из четырех фрагментов получены последовательности для 267 образцов из 45 популяций: Rh. ledebourii – 94; Rh. dauricum – 80, Rh. sichotense – 47 и 46 образцов Rh. mucronulatum. Сочетание изменчивости по 4 маркерным фрагментам позволило выявить 14 гаплотипов, сгруппированных в соответствии с их видовой принадлежностью и географической локализацией (рис. 1).
Наиболее полиморфны Rh. ledebourii (гаплотипы H1–H7), показатель изменчивости H варьировал от 0 до 0.56 и Rh. sichotense (H9–H11) показатель Н варьировал от 0 до 0.3. У Rh. mucronulatum присутствуют три гаплотипа (H8,H13, Н14), однако все популяции мономорфны. Rh. dauricum мономорфный, во всех исследованных выборках присутствует единственный гаплотип H7. Распределение гаплотипов показывает филогеографическую структуру. Родственные гаплотипы имеют тенденцию сосуществовать совместно (значение NST (0.985) выше, чем GST (0.897) (P<0.001). Иерархический анализ AMOVA показал значительную дифференциацию между всеми четырьмя видами: доля межгрупповой изменчивости с учетом родства гаплотипов составила 89.6%, без учета родства – 53.2%, что больше доли изменчивости, приходящейся на межпопуляционную составляющую внутри групп (с учетом родства гаплотипов и без – 8.9% и 36.5% соответственно).
Филогенетическое дерево гаплотипов (рис. 2) построено с помощью MrBayers ver. 3.2.3. (Huelsenbeck et al., 2011) с Rh. aureum в качестве внешней группы. Характер ветвления дерева поддерживает деление видов на две группы: восточно-сибирскую (Rh. ledebourii и Rh. dauricum) и приморскую (Rh. sichotense и Rh. mucronulatum). Парное сходство видов в этих группах отмечалось разными авторами на основании анатомо-морфологических параметров листа, а также по составу и содержанию некоторых химических веществ (Белоусов, 2000; Карпов, Каракулов, 2011). Заметим, что в данных группах у одного из видов присутствует признак листопадности (Rh. dauricum, Rh. mucronulatum), у двух других – полувечнозелености (Rh. ledebourii, Rh. sichotense).
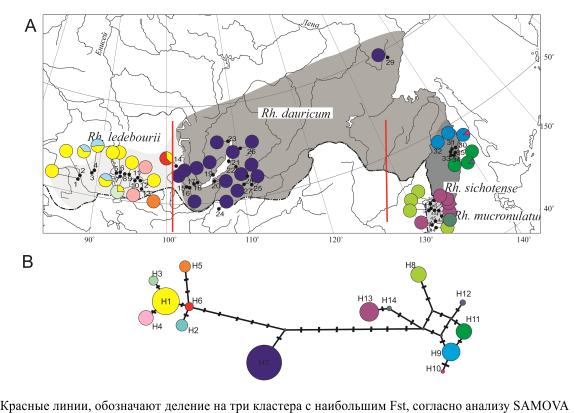
Рисунок 1 – А. Ареалы и месторасположение выборок, а также частоты хлоропластных гаплотипов изученных видов рододендронов Rh. ledebourii, Rh. dauricum, Rh. sichotense и Rh. mucronulatum в Сибири и на Дальнем Востоке; B. Дерево минимальных расстояний между хлоропластными гаплотипами (Н). Rh. ledebourii H1-H6, Rh. dauricum H7, Rh. sichotense H9-H12 и Rh. mucronulatum H8, H13, H14
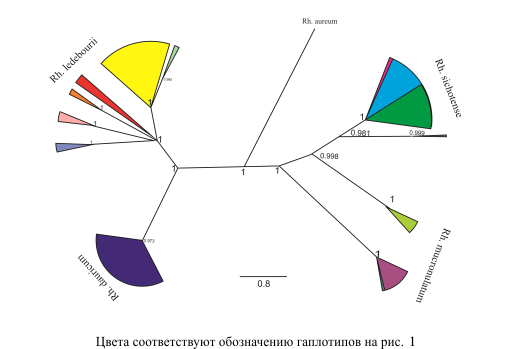
Рисунок 2 – Филогения популяций изученных видов рододендрона, в качестве аутгруппы использован образец Rh. aureum
Географические группы, выделенные с помощью SAMOVA, частично перекрываются с группами филогенетического дерева (рис. 1а, красные линии). Максимальное значение Fst наблюдалось при делении на три группы: I – Rh. ledebourii, II – Rh. dauricum, III – Rh. sishotense и Rh. mucronulatum. Выявленная степень дифференциации видов хорошо согласуется с их географическим положением и удаленностью друг от друга: Rh. ledebourii произрастает в горах Западного Саяна и на Алтае, Rh. dauricum – в горах Восточного Саяна, Северного Прибайкалья и Забайкалья, Rh. sichotense – в средней части Сихотэ-Алиня, а Rh. mucronulatum – на юге Приморья. Дивергенция их обусловлена филогенетической историей, сложившейся под влиянием климатических флуктуаций позднего плейстоцена. Виды с наибольшей изменчивостью Rh. ledebourii, Rh. sichotense и Rh. mucronulatum, вероятно, не изменяли значительно своей популяционной численности во время похолоданий в горных районах Южной Сибири и в Приморье, то есть на территориях известных в качестве рефугиумов, как по палеогеографическим, так и генетическим данным. Обособление Rh. dauricum обусловлено резким сокращением численности предковых популяций, и дальнейшей экспансией уже с обедненным полиморфизмом в результате прохождения через «бутылочное горлышко». Высокое генетическое разнообразие, наличие гаплотипов со схожими мутациями (рекомбинантых) свидетельствуют в пользу того, что виды Rh. ledebourii и Rh. sichotense можно считать более близкими к предковому виду, некогда распространенному на всей территории северо-восточной Сибири, а Rh. dauricum и Rh. mucronulatum их производными, соответственно. Этим логично объясняется близость в парах Rh. ledebourii – Rh. dauricum, Rh. sichotense – Rh. mucronulatum на основе анатомо-морфологических характеристик листа, состава вторичных метаболитов, а также переход от полувечнозелености к листопадности, как более молодому в эволюционном смысле признаку, связанному с адаптацией к похолоданию климата. Менее четкая дифференциация в приморской группе, а также промежуточное положение гаплотипа H7 (рис. 1b) указывают на возможные повторяющиеся в истории контакты Rh. dauricum, Rh. sichotense и Rh. mucronulatum (или их общего предшественника).