Впервые реконструирована история расселения можжевельника обыкновенного (Juniperus communis) в послеледниковые периоды на евразийском ареале. Выявленные закономерности не соответствуют ни одному известному сценарию постгляциальных расселений древесных видов растений
Для анализа генетической изменчивости и филогеографической истории одного из видов семейства кипарисовых – можжевельника обыкновенного (Juniperus communis sensu lato) впервые были использованы два фрагмента хлоропластной ДНК – trnT-trnF and 16s-trnA (42 популяции, 622 образца). В результате комбинации полиморфизма двух сайтов обнаружено 84 гаплотипа. Столь высокий уровень генетической изменчивости удивителен для хвойных и, возможно, связан с распространением этого вида перелетными птицами. Однако количество гаплотипов в разных популяциях сильно различалось. Так, в Якутской популяции из 8 собранных образцов обнаружено 7 разных гаплотипов, из них 4 редких, нигде более не встречающихся. В шведской популяции из 12 образцов – 7 гаплотипов, из них 2 редких. В то же время почти все 32 образца, собранные на Тянь-Шане, оказались принадлежащими к одному гаплотипу (рис. 1). Таким образом, показано, что наиболее высокий уровень внутрипопуляционного разнообразия существует в северных популяциях можжевельника, а южные популяции генетически обеднены.

Рисунок 1 – Внутрипопуляционное разнообразие He and π, основанное на 84 хлДНК гаплотипах Juniperus communis. Цвет и размер кругов относятся к значениям He и p, соответственно. He –белые круги: 0-0.49, серые круги - 0.55-0.83, черные круги – 0.85-0.96. p –маленькие круги: 0-0.028, средние круги – 0.031-0.051, большие круги – 0.056-0.18
Реконструкция филогенетических отношений выявила несколько высоко дивергентных генетических линий у можжевельника (GL1-GL4) и шесть SAMOVA групп (рис. 2). Самая большая клада GL3 включает популяции из Альп, Северной Европы и Сибири. Дифференциация между этими популяциями имеет средний уровень (Fst =28%) по сравнению с общим ареалом (Fst =76%), а Fs статистика имеет высокое отрицательное значение, указывая на популяционную экспансию. Некоторые гаплотипы внутри GL3 формируют субклады с ограниченным географическим распространением, подтверждая локальное происхождение мутаций и слабый генный поток.
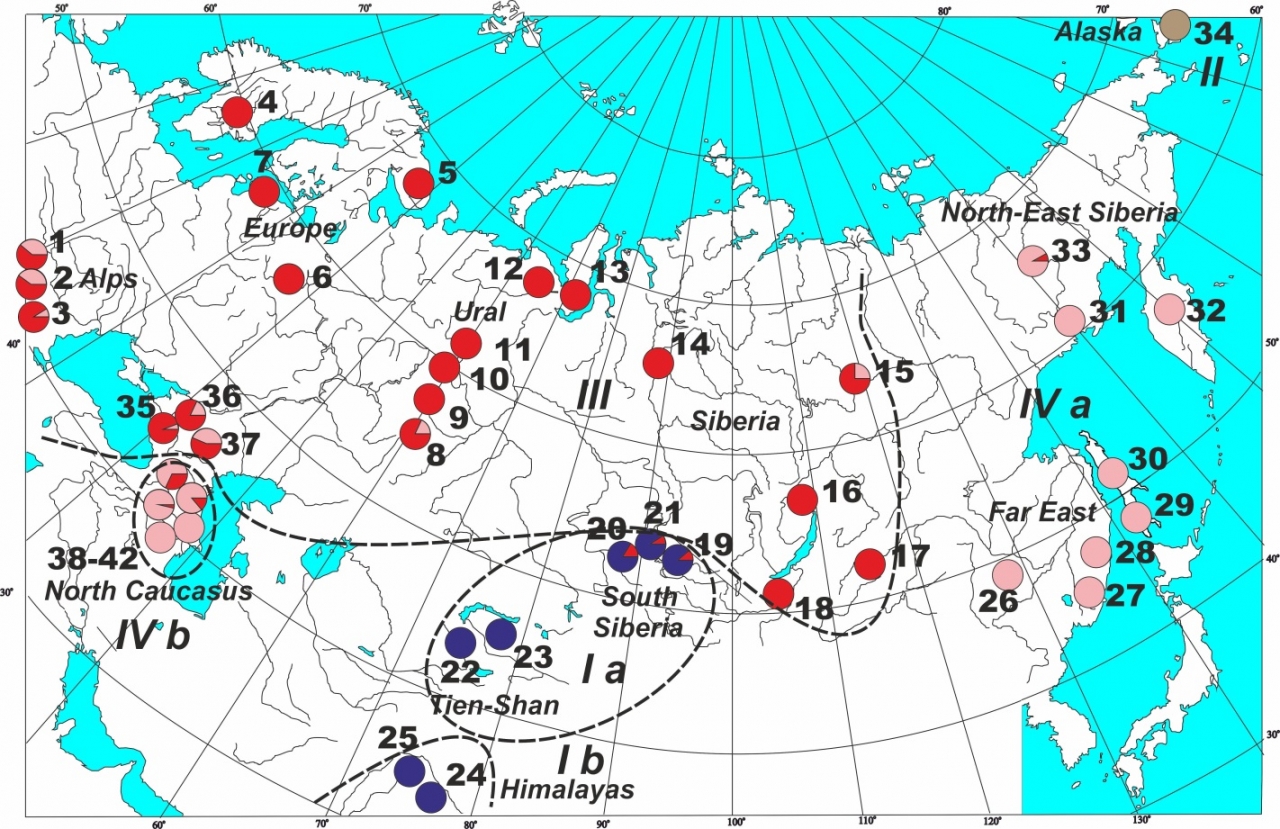
Рисунок 2 – Карта, показывающая расположение популяционных выборок Juniperus communis в Евразии и распределение главных генетических линий (GLs). Синий – GL1, коричневый – GL2, красный – GL3, розовый – GL4. Группы, выделенные согласно SAMOVA, обозначены римскими цифрами
Южные горные популяции можжевельника (восточная часть Северного Кавказа, Гималаи, Тянь-Шань и Южная Сибирь) и периферические популяции Дальнего Востока (GL1 и GL4) сильно дифференцированы от популяций остальной части ареала, что позволяет предположить, что они не вносили вклад в реколонизацию Северной Евразии. Скорее всего, колонизация Северной Европы и Сибири произошла из ограниченного источника (возможно, из Альп) еще до последнего ледникового максимума. Метод моделирования экологических ниш (рис. 3) не обнаружил значительного уменьшения ареала этого вида в течение последнего оледенения, поэтому можно допустить, что в последующие ледниковые эпохи можжевельник выживал в высоких широтах в микрорефугиумах, которые были вторичными центрами реколонизации.
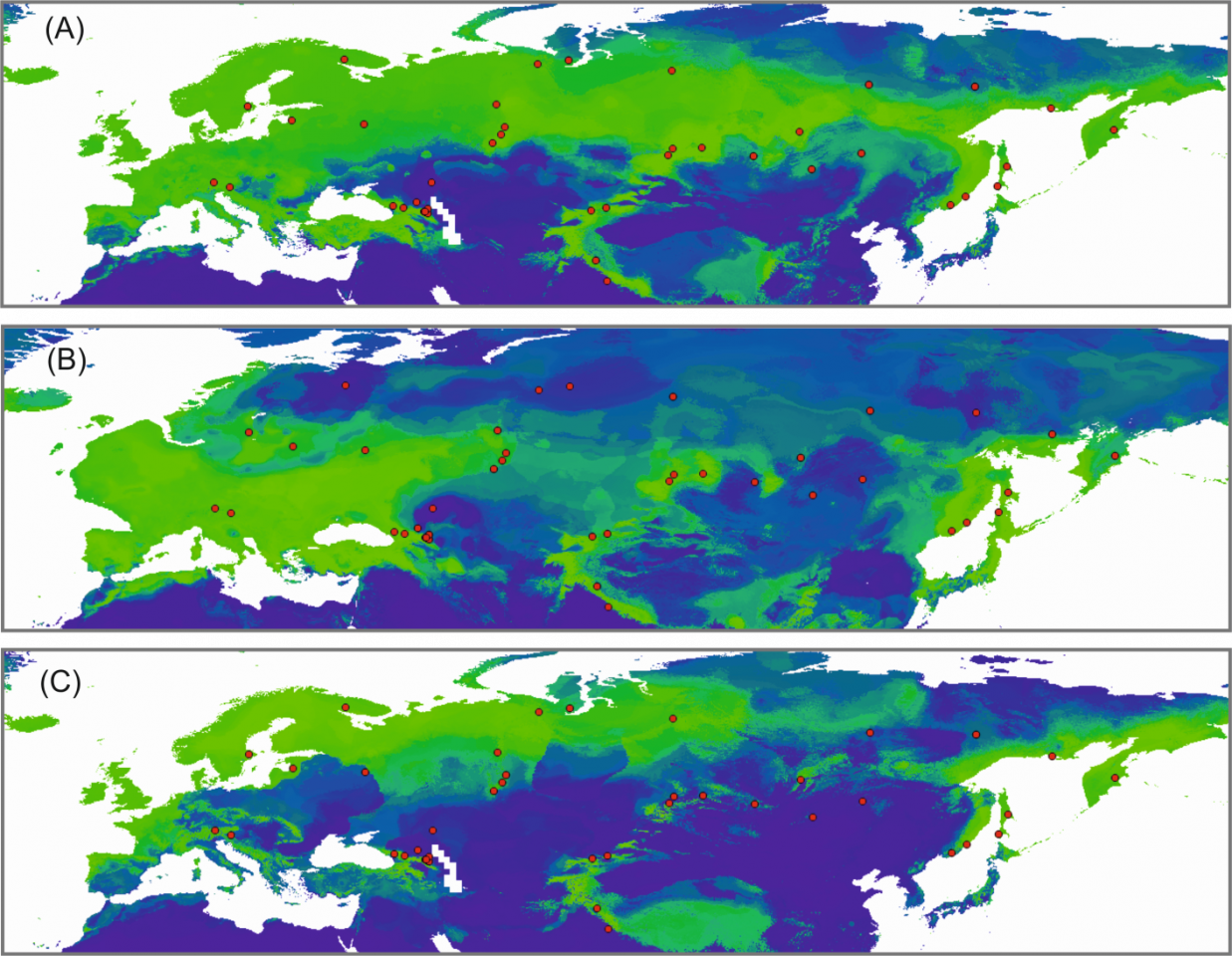
Рисунок 3 – . Моделирование экологических ниш у J. communis для (A) настоящего времени, (B) последний ледниковый максимум (21 000 лет) и (C) межледниковый период. Теплые цвета показывают высокую вероятность присутствия вида, в то время как более холодные цвета – более низкую вероятность