2014
Направление:
"Общая генетика"
На основе анализа географического распространения гаплотипов хлоропластной и митохондриальной ДНК впервые проведена филогенетическая реконструкция рода Abies. Установлено, что современные пихты берут начало в западной части Северной Америки, где наблюдается наиболее глубокая дифференциация отдельных линий и обитают представители базальных клад. В качестве пути миграций пихт между континентами можно рассматривать только берингийский мост.
Впервые для рода Abies использовались цитоплазматические маркеры с различным типом наследования и разной скоростью мутирования: наследуемая по отцовской линии хлоропластная ДНК (хпДНК) и наследуемая по материнской линии митохондриальная ДНК (мтДНК). Филогенетическое исследование включало 53 образца 37 видов пихт, представляющих основные линии эволюции рода, а также внешнюю группу Keteleeria davidiana. Методом секвенирования получены нуклеотидные последовательности пяти регионов хпДНК общей длиной 5580 пар нуклеотидов (п.н.) и трех фрагментов мтДНК общей длиной 5226 п.н. Изменчивость изученных хлоропластных фрагментов была существенно выше (обнаружено 101 информативных мутаций у Abies), чем изменчивость митохондриальных фрагментов (25 информативных мутаций). В результате получено наиболее полное на настоящий момент и имеющее наилучшее разрешение филогенетическое исследование Abies, основанное на сравнении данных двух геномов. Филогенетическая реконструкция рода на основе изменчивости хлоропластных фрагментов производилась с использованием методов максимальной парсимонии (MP) и байесовского подхода (BI). Топология MP- и BI- деревьев оказалась сходной и состоящей из шести основных групп видов Abies. Статистическая поддержка всех основных клад и большинства субклад в обеих топологиях была достаточно высокой, всего на BI- дереве представлено 23 клады с поддержкой (> 50%) (рис. 1).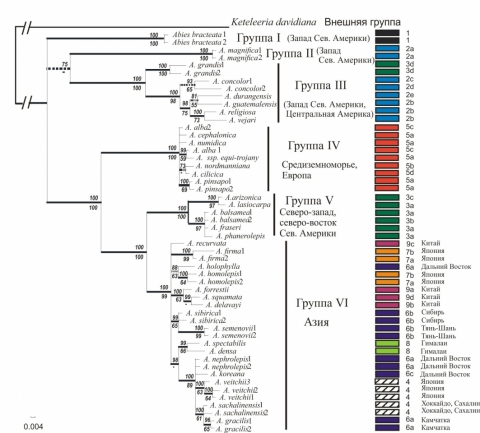 Над линиями приведена статистическая поддержка байесовским методом (значения апостериорной вероятности, умноженные на 100), под линиями – бутстрап - поддержка (MP, 1000 репликаций). Толстые горизонтальные линии обозначают поддержку более 95%, прерывистые линии – поддержку между 50 и 95%. Прочерк означает воспроизводимость данной клады в менее чем 50% реплик бутстрапа (MP). Шкала отражает число замен на нуклеотидную позицию. Приведены митотипы исследованных образцов Abies, наложенные на филогенетическое дерево
Рисунок 1 – Филогенетическое дерево Abies, построенное с помощью байесовского метода (BI, 50% консенсус), для 53 образцов Abies и Keteleeria на основе нуклеотидных последовательностей фрагментов хлоропластной ДНК общей длиной 5580 пар нуклеотидов
Для изучения взаимоотношений полученных митохондриальных гаплитопов (митотипов) был использован метод “median-joining network” (рис. 2). Сеть митотипов состояла из двух больших кластеров: кластер I, «американский», представлял американские виды и два азиатских, кластер II – все оставшиеся евроазиатские виды. Внутри двух кластеров 25 митотипов образовали 9 основных групп, в основном соответствующих кладам хпДНК филогении (рис. 1), однако их взаимоотношения оказались существенно отличными: принадлежность вида к группе митотипов в большей степени соответствовала его географическому распространению. Филогенетическая сеть митотипов имела высокий уровень гомоплазий и содержала циклы, которые свидетельствуют о рекомбинации как важном факторе эволюции мтДНК у Abies.
Над линиями приведена статистическая поддержка байесовским методом (значения апостериорной вероятности, умноженные на 100), под линиями – бутстрап - поддержка (MP, 1000 репликаций). Толстые горизонтальные линии обозначают поддержку более 95%, прерывистые линии – поддержку между 50 и 95%. Прочерк означает воспроизводимость данной клады в менее чем 50% реплик бутстрапа (MP). Шкала отражает число замен на нуклеотидную позицию. Приведены митотипы исследованных образцов Abies, наложенные на филогенетическое дерево
Рисунок 1 – Филогенетическое дерево Abies, построенное с помощью байесовского метода (BI, 50% консенсус), для 53 образцов Abies и Keteleeria на основе нуклеотидных последовательностей фрагментов хлоропластной ДНК общей длиной 5580 пар нуклеотидов
Для изучения взаимоотношений полученных митохондриальных гаплитопов (митотипов) был использован метод “median-joining network” (рис. 2). Сеть митотипов состояла из двух больших кластеров: кластер I, «американский», представлял американские виды и два азиатских, кластер II – все оставшиеся евроазиатские виды. Внутри двух кластеров 25 митотипов образовали 9 основных групп, в основном соответствующих кладам хпДНК филогении (рис. 1), однако их взаимоотношения оказались существенно отличными: принадлежность вида к группе митотипов в большей степени соответствовала его географическому распространению. Филогенетическая сеть митотипов имела высокий уровень гомоплазий и содержала циклы, которые свидетельствуют о рекомбинации как важном факторе эволюции мтДНК у Abies.
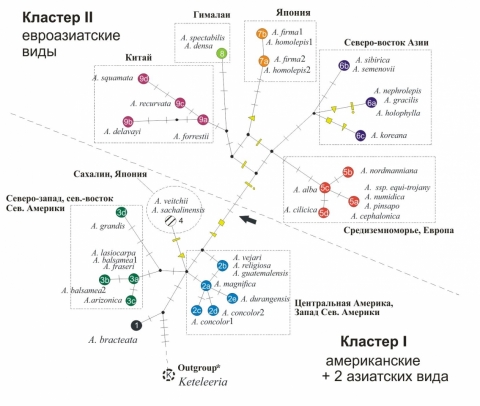 Группы митотипов, обозначенные разной заливкой, и клады, разделенные штриховой линией, обсуждаются в тексте. Черные поперечные линии обозначают мутации. Специальные значки обозначают некоторые мутации – гомоплазии. Черной стрелкой обозначена инсерция во фрагменте nad4(3/4) . При построении филогенетической сети не учитывались мутации, специфичные для внешней группы Keteleeria davidiana.
Рисунок 2 – Филогенетическая сеть митотипов Abies и Keteleeria, построенная на основе нуклеотидных последовательностей фрагментов мтДНК с помощью метода “median-joining network” (Bandelt et al., 1999)
Полученная филогенетическая реконструкция с оценками возраста событий дивергенции позволяет высказать ряд гипотез относительно истории рода Abies. Калибровки возраста узлов филогенетического дерева, проведенные на основании палеоботанических данных и на оценке скорости мутирования хлоропластной ДНК у Pinaceae, дали близкие результаты (рис. 3). Молекулярная датировка предполагает расхождение базальных клад современных Abies на границе между олигоценом и миоценом (HPD95% = 18–27 млн л.н.). Происхождение современных пихт берет начало в западной части Северной Америки, где наблюдается наиболее глубокая дифференциация отдельных линий Abies, и поскольку там обитают представители базальных клад – наиболее рано отделившаяся от основного дерева A. bracteata и, в равной степени, группа A. magnifica и родственных ей видов. Евразийские центры разнообразия пихт, безусловно, являются вторичными по отношению к североамериканским. В качестве пути миграций современных линий пихт между континентами можно рассматривать только берингийский мост. В конце олигоцена – начале миоцена произошла миграция Abies из Америки в Азию, и далее в Средиземноморье. Оценка возраста отделения европейско-средиземноморских пихт от азиатских соответствует миоцену. Во второй половине миоцена произошла обратная миграция из Азии в Америку, что соответствует образованию отдельной ветви «бореальных» американских видов (A. lasiocarpa, A. balsamea и др.) от азиатских видов, имевшим место в позднем миоцене. Дифференциация внутри большинства азиатских видов Abies происходила с конца миоцена, в течение плиоцена и плейстоцена.
Группы митотипов, обозначенные разной заливкой, и клады, разделенные штриховой линией, обсуждаются в тексте. Черные поперечные линии обозначают мутации. Специальные значки обозначают некоторые мутации – гомоплазии. Черной стрелкой обозначена инсерция во фрагменте nad4(3/4) . При построении филогенетической сети не учитывались мутации, специфичные для внешней группы Keteleeria davidiana.
Рисунок 2 – Филогенетическая сеть митотипов Abies и Keteleeria, построенная на основе нуклеотидных последовательностей фрагментов мтДНК с помощью метода “median-joining network” (Bandelt et al., 1999)
Полученная филогенетическая реконструкция с оценками возраста событий дивергенции позволяет высказать ряд гипотез относительно истории рода Abies. Калибровки возраста узлов филогенетического дерева, проведенные на основании палеоботанических данных и на оценке скорости мутирования хлоропластной ДНК у Pinaceae, дали близкие результаты (рис. 3). Молекулярная датировка предполагает расхождение базальных клад современных Abies на границе между олигоценом и миоценом (HPD95% = 18–27 млн л.н.). Происхождение современных пихт берет начало в западной части Северной Америки, где наблюдается наиболее глубокая дифференциация отдельных линий Abies, и поскольку там обитают представители базальных клад – наиболее рано отделившаяся от основного дерева A. bracteata и, в равной степени, группа A. magnifica и родственных ей видов. Евразийские центры разнообразия пихт, безусловно, являются вторичными по отношению к североамериканским. В качестве пути миграций современных линий пихт между континентами можно рассматривать только берингийский мост. В конце олигоцена – начале миоцена произошла миграция Abies из Америки в Азию, и далее в Средиземноморье. Оценка возраста отделения европейско-средиземноморских пихт от азиатских соответствует миоцену. Во второй половине миоцена произошла обратная миграция из Азии в Америку, что соответствует образованию отдельной ветви «бореальных» американских видов (A. lasiocarpa, A. balsamea и др.) от азиатских видов, имевшим место в позднем миоцене. Дифференциация внутри большинства азиатских видов Abies происходила с конца миоцена, в течение плиоцена и плейстоцена.
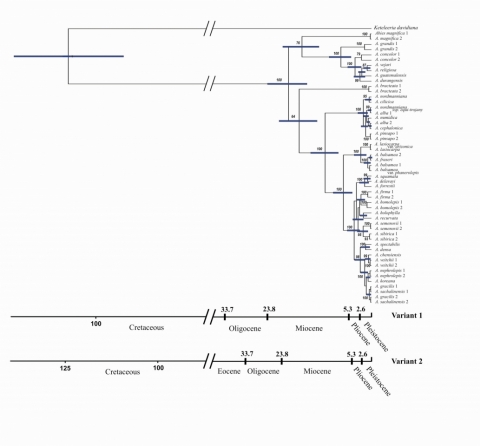 Прямоугольники при узлах обозначают соответствующие варианту калибровки 1 доверительные 95%-ные интервалы времени дивергенции (HPD95%, highest posterior density – наибольшая постериорная плотность)
Рисунок 3– Байесовская хронограмма филогении видов пихт, полученная в программе BEAST. Оценки времени дивергенции проведены с помощью двух вариантов калибровки. Вариант 1 - калибровка основана на палеоботанических данных с оценкой расхождения родов Abies и Keteleeria (100.4–113.8 млн. лет назад, Kremp, 1967; Xiang et al., 2007). Вариант 2 - калибровка основана на оценке скорости накопления молчащих замен в хлоропластном геноме видов Pinaceae, 0.22•10-9 site-1 year-1 (Willyard et al., 2007)
Возраст диверсификации внутри групп средиземноморских, «бореальных» американских, мексиканских видов Abies оценивается как плиоцен-плейстоцен (рис. 3). При значительном обособлении средиземноморских пихт от остальных групп, сами они оказываются весьма слабо дифференцированными. Эволюционную близость всех средиземноморских видов между собой можно объяснить «бутылочным горлышком», связанным с происходившим в позднем миоцене (5,6 млн. лет назад) в бассейне Средиземного моря мессинским кризисом солености, приведшим к сильной аридизации окружающих территорий, или же с другими неблагоприятными климатическими событиями, имевшими место в плиоцене. Дивергенция в группе мексиканских видов пихт произошла в плио- и плейстоцене, что соответствует возрасту наиболее ранних находок пихты в Мексике и отражает относительно недавнюю историю проникновения пихт в местные горные системы. Дифференциация «бореальных» американских пихт на западную (A. lasiocarpa) и восточную (A. balsamea) и другие виды восточного побережья Северной Америки) клады произошла в плейстоцене и, вероятно, в связи с начавшейся эпохой оледенений и связанным с ней биогеографическим разрывом между тихоокеанской и атлантической частями бореальной биоты Северной Америки.
Обнаруженные случаи неконгруэнтности между филогенией хпДНК и филогенетической сетью митотипов предполагают захват мтДНК при интрогрессивной гибридизации видов. Согласно проведенным сравнительным молекулярно-филогенетическим исследованиям, события расселения между Северной Америкой и Азией через берингийский мост могли быть неоднократными и происходить в обоих направлениях. Несоответствие между “евроазиатской” хпДНК и “американской” мтДНК “бореальных” американских видов, вероятно, является следствием захвата ими мтДНК аборигенных американских видов при их проникновении в Америку из Азии. С другой стороны, митотипы, родственные «американскому» кластеру мтДНК, были обнаружены у некоторых азиатских видов, ареалы которых расположены в северо-западной части тихоокеанской арки, на линии возможных миграций между двумя континентами. Можно заключить, что ретикулярная эволюция, вызванная межвидовым генным потоком мтДНК и рекомбинантным преобразованием родительских линий мтДНК, играет значительную роль в эволюционной истории Abies.
Прямоугольники при узлах обозначают соответствующие варианту калибровки 1 доверительные 95%-ные интервалы времени дивергенции (HPD95%, highest posterior density – наибольшая постериорная плотность)
Рисунок 3– Байесовская хронограмма филогении видов пихт, полученная в программе BEAST. Оценки времени дивергенции проведены с помощью двух вариантов калибровки. Вариант 1 - калибровка основана на палеоботанических данных с оценкой расхождения родов Abies и Keteleeria (100.4–113.8 млн. лет назад, Kremp, 1967; Xiang et al., 2007). Вариант 2 - калибровка основана на оценке скорости накопления молчащих замен в хлоропластном геноме видов Pinaceae, 0.22•10-9 site-1 year-1 (Willyard et al., 2007)
Возраст диверсификации внутри групп средиземноморских, «бореальных» американских, мексиканских видов Abies оценивается как плиоцен-плейстоцен (рис. 3). При значительном обособлении средиземноморских пихт от остальных групп, сами они оказываются весьма слабо дифференцированными. Эволюционную близость всех средиземноморских видов между собой можно объяснить «бутылочным горлышком», связанным с происходившим в позднем миоцене (5,6 млн. лет назад) в бассейне Средиземного моря мессинским кризисом солености, приведшим к сильной аридизации окружающих территорий, или же с другими неблагоприятными климатическими событиями, имевшими место в плиоцене. Дивергенция в группе мексиканских видов пихт произошла в плио- и плейстоцене, что соответствует возрасту наиболее ранних находок пихты в Мексике и отражает относительно недавнюю историю проникновения пихт в местные горные системы. Дифференциация «бореальных» американских пихт на западную (A. lasiocarpa) и восточную (A. balsamea) и другие виды восточного побережья Северной Америки) клады произошла в плейстоцене и, вероятно, в связи с начавшейся эпохой оледенений и связанным с ней биогеографическим разрывом между тихоокеанской и атлантической частями бореальной биоты Северной Америки.
Обнаруженные случаи неконгруэнтности между филогенией хпДНК и филогенетической сетью митотипов предполагают захват мтДНК при интрогрессивной гибридизации видов. Согласно проведенным сравнительным молекулярно-филогенетическим исследованиям, события расселения между Северной Америкой и Азией через берингийский мост могли быть неоднократными и происходить в обоих направлениях. Несоответствие между “евроазиатской” хпДНК и “американской” мтДНК “бореальных” американских видов, вероятно, является следствием захвата ими мтДНК аборигенных американских видов при их проникновении в Америку из Азии. С другой стороны, митотипы, родственные «американскому» кластеру мтДНК, были обнаружены у некоторых азиатских видов, ареалы которых расположены в северо-западной части тихоокеанской арки, на линии возможных миграций между двумя континентами. Можно заключить, что ретикулярная эволюция, вызванная межвидовым генным потоком мтДНК и рекомбинантным преобразованием родительских линий мтДНК, играет значительную роль в эволюционной истории Abies.
Исполнители: